Fluctuating selection study in PNAS!
The result of two years of hard work with Luis-Miguel Chevin during my last post-doc in CEFE (Montpellier, France) has just been published in Proceedings of the National Academy of Sciences of the United States of the America (or PNAS, for the lazy)! Here is the final, accepted version of the manuscript.
A bit of context
This study is part of Luis-Miguel’s ERC project on the study of fluctuating selection. It notably included very cool stuff on experimental evolution on a halophile algae, Dunaliella salina, involving random fluctuations of the salinity and… a robot! Check this out here. The part of the project I worked on was more focused on studying fluctuating selection in wild populations, in order to find more about its prevalence and importance in a natural setting.
Fluctuating selection, what?
But what is fluctuating selection? It’s when the characteristics of natural selection (via the individual fitness) fluctuates over time. This is expected because the environment itself randomly changes over time (think about weather for example). It is probably easiest to understand when “time” means “year” for example: you would expect the selective pressure to not be exactly identical from a year to another, as some years are warmer, or dryer, or with a different timing of first frosts, than others. In our project, we referred to the consequences of such variations over time on the individual fitness function, relating the phenotypic trait (the actual, observed characteristics of the individuals) values to the individual fitness, as “fluctuating selection”.
A complication is that the phenotypic trait values can also change with the environment, a phenomenon biologists call “phenotypic plasticity”. If plasticity is perfect, then fluctuations of the fitness function as described above should result in no variation of the selection from a year to another, because the phenotype would have changed accordingly. So, we also studied how much of these variation in the selection characteristics translate into variations in the actual, observable selection in the wild populations. OK, this is becoming confused, this is all probably a bit clearer on the following cartoon:
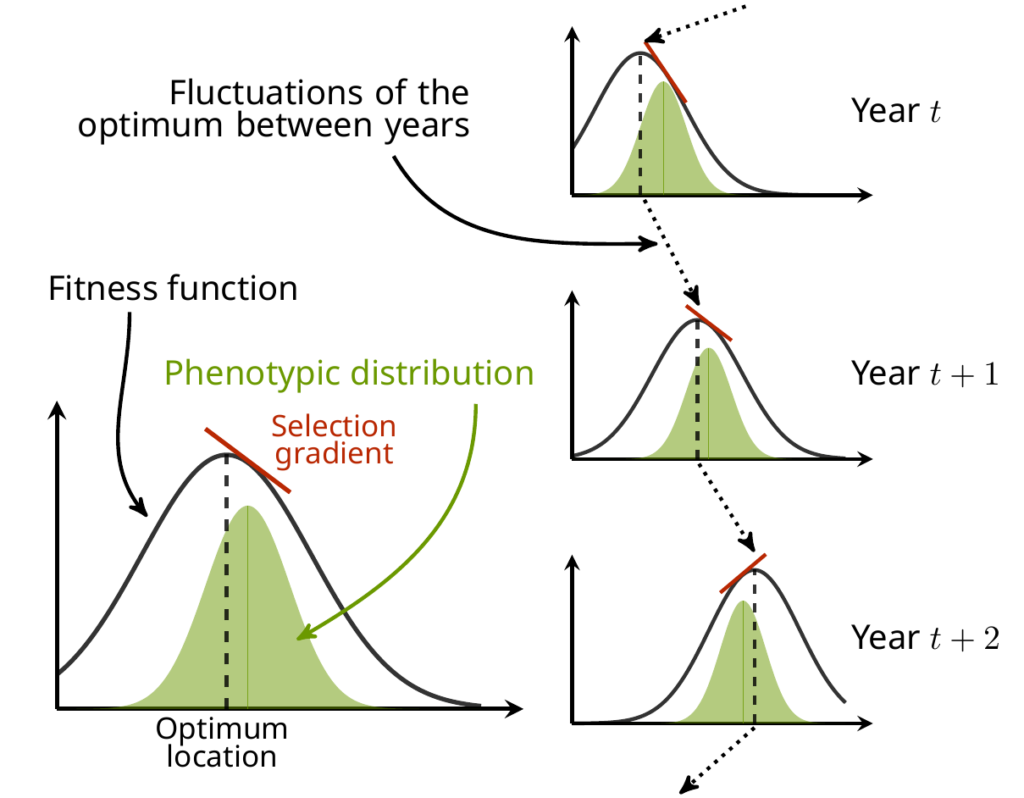
The method
To do all this, with the help of Anne Charmantier, we nicely asked a lot of people if they wanted to contribute their long-term data of wild populations to our project. We decided to focus on “breeding phenology”, or timing of reproduction, or lay date for birds, or parturition date for mammals… Well, basically, when individuals decide to reproduce, as measured by… when they reproduce. Many (most!) of the people of people we asked answered very positively!
In the end, we gathered 39 datasets on 21 species of birds and mammals (those two taxa make the bunch of long-term individual surveys for many reasons). We designed a statistical model corresponding to the fluctuating optimum as shown above and apply this same model to each datasets (we also applied models without an optimum, but still with fluctuations of the fitness parameters, like the slope, and the same models without fluctuations: we explored a few things, don’t worry).
What we found!
We found that the model including an optimum was a good description of the data behaviour for the most part, and that, optimum or not, there was clear signs of fluctuations in the parameters of our fitness functions. This translated into smaller, but clearly present variations in actual selection, which would mean that phenotypic plasticity is there, but certainly not perfectly tracking the fluctuations. We also confirmed a well-known results for birds: there was a consistent selection for earlier lay dates. This phenomenon is described here and here, for example (and here, shameless self-promotion).
If you think about it, it’s weird, this thing about birds: we have consistent selection for earlier breeding, and yet, the fluctuations of the optimum do not entirely translate into variations in actual, observed selection, which would point to a bit of phenotypic plasticity, able at least to some extent “track” the fluctuations of the optimum, right? Well, you can have both at the same time if… the phenotypic mean tracks the optimum, but “from afar”. OK, it’s hard to explain, it’s better visualised with an animation of the fluctuations of both the optimum and phenotypic mean over the years:
We wanted to confirm this tracking was the result of phenotypic plasticity. This is when we remembered: we also have the individual ID of each breeding female, so we can find individuals that bred on two consecutive years, their change in phenotype could only arise through phenotypic plasticity, right? So, we did that, computed the change due to “phenotypic plasticity alone” and compared it to the change in the optimum location and found a significant correlation for 4 datasets, and an overall significance for all bird datasets put together! So, this seems to be phenotypic plasticity, alright.
OK, so what?
Well, this is interesting for a few reasons. First, we found that fluctuations in the fitness characteristics are very prevalent, and although they only partly translate into variations in actual, observed selection, this is because phenotypic plasticity itself evolved to respond to those fluctuations! Second, we provided “meta-estimates” (averaged over the datasets) for models with a fluctuating optimum, which would be very useful for theoreticians in the future. Third, there was this very intriguing result that birds, in many populations, are tracking fluctuations of the optimum, but “from afar”. I personally think this would mean there is a constraint in this selection for earlier breeding we do not understand. It could be that a scenario proposed by Price, Kirkpatrick & Arnold (1998), whereby the selection for earlier breeding is a sort of “illusion”, even at the evolutionary equilibrium, due to inter-individual competition for breeding resources (like nest places), might have some real value in explaining this result.